リン酸仮説:7.糖・有機酸代謝の発生
高エネルギーリン酸化合物産生システムの発生(== 現代の解糖系やペントースリン酸サイクル)の起源を論じる.
このシステムに含まれる酵素活性には,酸化還元活性,アルドラーゼ活性,イソメラーゼ活性が含まれる.
これらの反応は,ビルディングブロックからコンポーネントが生成する脱水縮合反応とは別物である.
酸化還元活性の補酵素 は NAD(P)H+ や FAD(P)H2 なので,少なくとも互換性のある分子が形成されていなくてはならい.
リン酸仮説ではビルディングブロック(糖,リン酸,核酸塩基,およびアミノ酸)を組み合わせて生命の発生を論じている.
ビルディングブロックの量が不変であるならば,その最小濃度のものがプロトセル(あるは初期の細胞)の量を規定してしまう.
不足分は中間代謝で供給されなければならない,というわけである.
目次(ページ内リンク)
解糖系とペントースリン酸サイクルとの共通部分
糖・有機酸代謝の発生
その後の進化
解糖系とペントースリン酸サイクルとの共通部分
糖のリン酸エステルをインキュベートするだけで,解糖系やペントースリン酸サイクルの主要反応は,非酵素的に進行する.
また,Fe(II) 等の遷移金属存在下では,それらの反応が促進される.
これらの反応は化学進化の初期から利用されたのであろう.
現代の解糖系とペントースリン酸サイクルを参照する.下記の特徴を有する.
- 解糖系とペントースリン酸サイクルには共通する部分がある
- 共通部分の基質は,糖または有機酸のリン酸誘導体である
解糖系
解糖系を「降りていく」方向に列挙し,酵素その他を付け加えると下のようになる.
フルクトース-6-リン酸
ATP ↓ 6-phosphofructokinase(EC 2.7.1.11) + ADP
フルクトース-1,6-ビスリン酸
↓ Fructose-bisphosphate aldolase(EC 4.1.2.13)
ジヒドロキシアセトンリン酸 + グリセルアルデヒド-3-リン酸
NAD+ + Pi↓ Glyceraldehyde-3-phosphate dehydrogenase(EC 1.2.1.12) + NADH
1,3-ビスホスホグリセリン酸
ADP↓ Phosphoglycerate kinase(EC 2.7.2.3) + ATP
3-ホスホグリセリン酸
ペントースリン酸サイクル
一方,ペントースリン酸サイクルでは同じ経路を「登ってゆく」.
フルクトース-6-リン酸
H2O ↑ Fructose-bisphosphatase(EC 3.1.3.11) + Pi
フルクトース-1,6-ビスリン酸
↑ Fructose-bisphosphate aldolase(EC 4.1.2.13)
ジヒドロキシアセトンリン酸 + グリセルアルデヒド-3-リン酸
NADPH ↑ Glyceraldehyde-3-phosphate dehydrogenase(EC 1.2.1.13) + NADP+ + Pi
1,3-ビスホスホグリセリン酸
ATP ↑ Phosphoglycerate kinase(EC 2.7.2.3) + ADP
3-ホスホグリセリン酸
TCA サイクルとの関係
解糖系の「共通部分」から TCA サイクルからオキサロ酢酸に至るまでに,3 〜 4 段階ある.
ホスホエノールピルビン酸の利用については,炭酸を結合するか脱炭酸するかの経路がある.
TCA サイクルのコンポーネントであるオキサロ酢酸になると,リン酸誘導体ではなくなる.
これは TCA サイクルが,より遅い時代に発生したことを意味すると解釈しておく.
3-ホスホグリセリン酸
↓ Phosphoglycerate mutase(EC 5.4.2.12)
2-ホスホグリセリン酸
↓ Phosphopyruvate hydratase(EC 4.2.1.11) + H2O
ホスホエノールピルビン酸
H2O + HCO3 ↓ Phosphoenolpyruvate carboxylase(EC 4.1.1.31) + Pi
オキサロ酢酸(TCA サイクルのコンポーネント)
あるいは
ADP ↓ Pyruvate kinase(EC 2.7.1.40) + ATP
ピルビン酸(脱炭酸してオキサロ酢酸となると,TCA サイクルのコンポーネント)
糖・有機酸代謝の発生
上の情報を踏まえて,糖・有機酸代謝が発生した道筋を考える.
- 化学進化のごく初期には,高エネルギーリン酸結合として,環境に存在するポリリン酸を利用していた
- ポリリン酸が枯渇するようになると,有機酸のリン酸誘導体(1,3-ビスホスホグリセリン酸エステル,ホスホエノールピルビン酸)を利用してポリリン酸を合成するようになった
- これらが枯渇してくると,似た分子からこれらのリン酸誘導体を酵素的に変換して,ポリリン酸を合成するようになった
解糖系の高エネルギーリン酸化合物
高エネルギーリン酸化合物は,プロトポリマーやプロトリン脂質のリン酸エステルの形成に関与したであろう.
特に,PRPP 経由の N-グリコシド結合の生成反応(その互換反応)に必要だったと考える.
現在の解糖系その他を眺めて,高エネルギーリン酸化合物をリストアップしてみると,
- リン酸ーリン酸無水物(== アデノシン三リン酸)
- リン酸ーカルボン酸無水物(== 1,3-ビスホスホグリセリン酸)
- リン酸-エノールエステル(== ホスホエノールピルビン酸)
化学進化の当初は,環境に存在していたと期待するポリリン酸などの高エネルギーリン酸化合物を利用していたはずである.
最終的に,上記の化合物を利用するようになったということだ.
最初の糖・有機酸代謝
環境の高エネルギーリン酸化合物が不足するような場合を想定すると,高エネルギーリン酸結合を保存できると有利である.
例えば,結合エネルギーの一部を代償にして,リン酸-エノールエステルをリン酸ーリン酸無水物に変換して保存する.
すなわち,
ホスホエノールピルビン酸
X ↓ X-P(何らかの形で高エネルギーリン酸結合を保存.以下では,↓ ATP と記す)
ピルビン酸
代謝カスケードの発達 その 1
ホスホエノールピルビン酸が枯渇してくると,何らかの方法で調達せねばならない.
2-ホスホグリセリン酸
↓
ホスホエノールピルビン酸
↓ ATP
ピルビン酸
代謝カスケードの発達 その 2
上と同じ理由で,カスケードが延長する.
解糖系やペントースリン酸サイクルの起源につながるように描いた.
また,最初にリブロースを置いたのは,リボースの起源を想定しているからである.
リブロース-1,5-ビスリン酸
↓
3-ホスホグリセリン酸
↓
2-ホスホグリセリン酸
↓
ホスホエノールピルビン酸
↓ ATP
ピルビン酸
解糖系の起源
3-ホスホグリセリン酸調達の別経路.
ペントースリン酸サイクルの起源を論じる都合上,フルクトース-1,6-リン酸を出発物質とする,
フルクトース-1,6-ビスリン酸
↓
ジヒドロキシアセトンリン酸 + グリセルアルデヒド-3-リン酸
↓
1,3-ビスホスホグリセリン酸
↓ ATP
3-ホスホグリセリン酸
↓
2-ホスホグリセリン酸
↓
ホスホエノールピルビン酸
↓ ATP
ピルビン酸
ペントースリン酸サイクルの起源
上のフルクトース-1,6-ビスリン酸 〜 3-ホスホグリセリン酸は平衡反応である.
したがって,フルクトース-1,6-ビスリン酸からリブロース-1,5-ビスリン酸への変換が可能であれば,ペントースリン酸サイクルとなる.
3-ホスホグリセリン酸
↓
1,3-ビスホスホグリセリン酸
↓
ジヒドロキシアセトンリン酸 + グリセルアルデヒド-3-リン酸
↓
フルクトース-1,6-ビスリン酸
↓
: 多段階のカスケード.このなかに,リボース-5-リン酸 → リブロース-5-リン酸の反応により,リボースが流入する部位がある
↓
リブロース-1,5-ビスリン酸
↓ ここで炭酸同化.光合成生物の分岐ポイントかも
3-ホスホグリセリン酸
リボースとリブロースの相互変換
上で想定している代謝カスケードのリブロース-1,5-ビスリン酸をホスファターゼとイソメラーゼで処理すれば,リボース-5-リン酸が生成する.
この逆反応は,リボース-5-リン酸がペントースリン酸サイクルに流入する経路である.
リブロース-1,5-ビス-リン酸
↓ リブロースビスホスファターゼ(未調査のため仮称.おそらく平衡反応)
リブロース-5-リン酸
↓ リボース-5-リン酸イソメラーゼ(未調査のため仮称.おそらく平衡反応)
リボース-5-リン酸
なお,リブロース-1,5-ビスリン酸とリボース-5-リン酸を,それぞれフルクトース-1,6-ビスリン酸とグルコース-6-リン酸に置き換えると解糖系の初期段階となる(流れは逆).
仮想的な代謝マップ
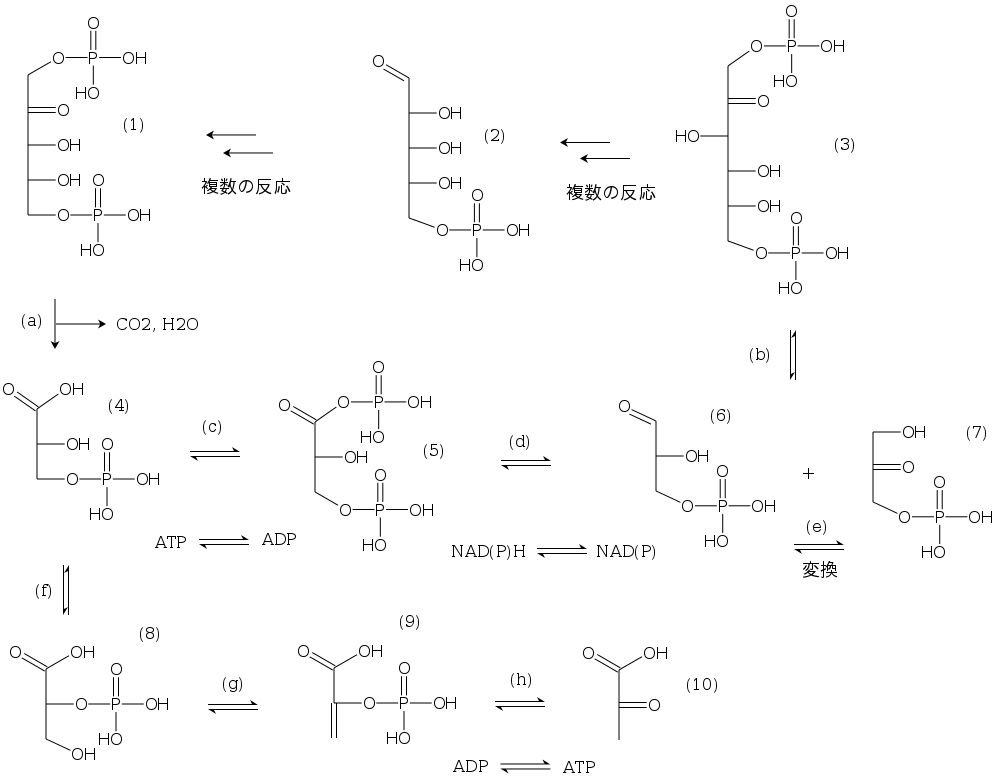
以上の考察を代謝マップにまとめた.
この部分は,TCA サイクルの近くである.
ペントースリン酸サイクル内ではリブロースとリボースはイソメラーセにより相互変換可能なので,リボースも記しておいた.
(1) リブロース-1,5-ビスリン酸
(2) リボース-5-リン酸
(3) フルクトース-1,6-ビスリン酸
(4) 3-ホスホグリセリン酸
(5) 1,3-ビスホスホグリセリン酸
(6) グリセルアルデヒド-3-リン酸
(7) ジヒドロキシアセトンリン酸
(8) 2-ホスホグリセリン酸
(9) ホスホエノールピルビン酸
(10) ピルビン酸
(a) Ribulose-bisphosphate carboxylase
(b) Fructose-bisphosphate aldolase
(c) Phosphoglycerate kinase
(d) Glyceraldehyde-3-phosphate dehydrogenase
(e) Triose-phosphate isomerase
(f) Phosphoglycerate mutase
(g) Phosphopyruvate hydratase
(h) Pyruvate kinase
解糖系は,3 → 6 + 7 → 5 → 4 → 8 → 9 → 10.
ペントースリン酸サイクルは,1 → 4 → 5 → 6 + 7 → 3 → 2 → 1.
煩雑になるので,この仮想的な代謝マップには FAD/FADH2 は記していない.
FADH2 の還元力は,NAD(P)+ の還元に利用される.
生成した FAD を還元する物質はシステイン(現在はリポ酸),ジスルフィド結合を還元する物質は Fe2+ あるいは Cu+ としておく.
その後の進化
煩雑になるので,高エネルギーリン酸結合の供給源を ATP と表す.
リン酸仮説では,N-グリコシド結合を形成するためには,リボースのヘミアセタール基がリン酸化されなければならないと仮定している.
ATP の導入により,N-グリコシドの生成と解糖系やペントースリン酸サイクルとが連結される.
- 有機酸-リン酸 + ADP ⇔ 有機酸 + ATP
- リボース + ATP → リボース 1-リン酸 + ADP
- リボース 1-リン酸 + アミノ化合物 → N-グリコシド + Pi
この経路を変異させると,CoA,GDP-糖,UDP-糖,CDP-コリン,PAPS(3’−ホスホアデノシン 5'−ホスホ硫酸)などの導入が可能かもしれない.